生体分子内あるいは生体分子間で働いている弱い相互作用としては、水素結合、van
der Waals力、疎水性相互作用などがある。水素結合はvan der
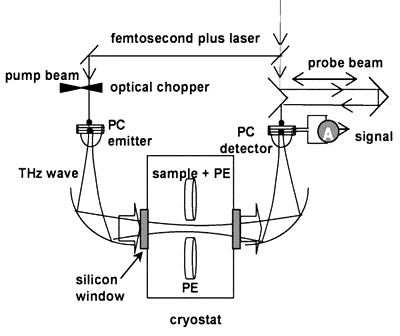
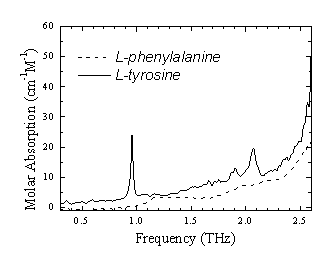
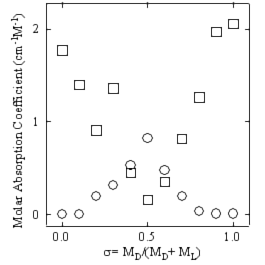
Waals力あるいは疎水性相互作用くらべると比較的強い相互作用であり、また生体分子の機能と構造に関連してさまざまな局面で重要な働きをしている。例えばDNAの二重らせんの形成はアデニン(A)とチミン(T)およびグアニン(G)とシトシン(C)の核酸のペアがそれぞれ二重及び三重の水素結合により結合することで形成されている。またタンパク質が目的とする機能を発揮するためにはその機能に対応する適当な三次元構造をとることが必要であるが、脱水状態ではそのような三次元構造をとることができない。すなわち水分子が水素結合を通してタンパク質分子と水和し、介添え役のような働きをすることで、その構造が維持されている。フェムト秒レーザーを用いてピコ秒電磁波パルスを発生させ、かつその電界をサンプリング検出するテラヘルツ時間領域分光法(THz-TDS)は、従来困難であった低周波数領域の吸収スペクトルを高い信号雑音比で測定することを可能にした。THz-TDSは、生体分子に働いている弱い相互作用を探る手段として有望である。
生体分子の低振動数モードすなわちTHz帯の振動モードは、THz-TDSの出現により、ようやく光があたり始めた状況と言える。今後THz-TDSが生体関連分子の機能及び構造研究にとって有用な分光手法となるためには、観測された振動モードと分子構造あるいは分子結晶構造との相関、及び水素結合やvan
der
Waals相互作用などとの関係の一般則を確立することが大きな課題である。これには、すでに確立されている中赤外域での手法が良いガイドとなると思われる。また、アミノ酸や糖類のような分子量の小さい分子だけではなく、タンパク質のようなマクロ分子の分光・分析をどのように行っていくかということも大きな課題である。タンパク質の場合、その構造は水和した水分子との相互作用を切り離して考えることができず、水溶液中あるいは水分を十分に制御した環境下でTHz吸収スペクトルを測定する必要がある。
A. G. Markelz, A. Roitberg and E.
J. Heilweil, “Pulsed terahertz spectroscopy of
DNA, bovine serum albumin and collagen between
0.1 and 2.0 THz”, Chem. Phys. Lett. Vol. 320, 42
(2000)
(2)
M. Brucherseifer, M. Nagel, P.
Haring Bolivar, H. Kurz, A. Bosserhoff and R.
Büttner, “Label-free probing of the binding
state of DNA by time-domain terahertz sensing”,
Appl. Phys. Vol. 77, 4049 (2000).
(3)
M. Nagel, P. Haring Bolivar, M.
Brucherseifer, and H. Kurz, A. Bosserhoff and R.
Büttner, “Integrated THz technology for
label-free genetic diagnostics”,Appl. Phys.
Lett. Vol.80, 154 (2002).
(4)
M. Walther, B. Fischer, M. Schall,
H. Helm, P. Uhd Jepsen, “Far-infrared
vibrational spectra of all-trans, 9-cis and
13-cis retinal measured by THz time-domain
spectroscopy”, Chem. Phys. Lett. Vol. 332, 389
(2000).
(5)
M. Tani, S. Matsuura, and K.
Sakai, “Emission Characteristics of
Photoconductive Antennas Based on
Low-Temperature-Grown GaAs and Semi-Insulating
GaAs,” Appl. Opt. Vol.36, 7853 (1997).
(6)
M.Hangyo, T. Nagashima and S.
Nashima, “Spectroscopy by pulsed terahertz
radiation”, Meas. Sci. Technol. Vol.13,1727,
(2002).
(7)
B.J. Johnson and G. H. Miller,
“Archaeological applications of amino acid
racemization.” Archaeolometry, Vol.39, No. 2,
pp.265-287 (1997).